Footnotes:
1In chimpanzees, adult males are generally considered to be more social than females (for details, check out the references at the bottom of this footnote), however, we don’t know if this sex difference is present from birth or emerges at some point during development. As play is the most common social interaction among young chimpanzees, that is one type of behavior that we can focus on to examine sex differences. As I analyze my dissertation data I’ll be trying to answer questions like: are males more playful than females? Do they initiate play more often? Spend more time playing? If so, how early does this sex difference emerge and how do general patterns of playfulness change with age? Do males and females play differently? Do they use different play styles? Focus on different partners? Do they use different play styles in response to the partners that they choose? (1) Goodall, J. (1986). The chimpanzees of Gombe: Patterns of behavior. (2) Wrangham, R. W. (1980). An ecological model of female-bonded primate groups. Behaviour, 75(3), 262-300. (3) Wrangham, Richard W. "Why are male chimpanzees more gregarious than mothers? A scramble competition hypothesis." Primate males: causes and consequences of variation in group composition. Cambridge University Press, Cambridge (2000): 248-258. (4) Otali, E., & Gilchrist, J. S. (2006). Why chimpanzee (Pan troglodytes schweinfurthii) mothers are less gregarious than nonmothers and males: the infant safety hypothesis. Behavioral Ecology and Sociobiology, 59(4), 561-570. (5) Lehmann, J., & Boesch, C. (2009). Sociality of the dispersing sex: the nature of social bonds in West African female chimpanzees, Pan troglodytes. Animal Behaviour, 77(2), 377-387.  Before we get started… Gender is a human cultural construct that ties up a lot of baggage linked to cultural definitions of the way that men and women should behave, rules about how they should interact among themselves and with each other, etc. However, we have no evidence that chimpanzees from differing communities exhibit culturally-based gender-roles. We also have no evidence that behavioral sex differences among chimpanzees are actively reinforced or shaped by other community members. For these reasons, we leave gender out of the chimpanzee conversation. Instead of using those loaded terms, we refer to males and females and discuss sex instead of gender. This is why I have edited the original wording of some of the questions. :)  Young Tambara pant-grunting at all the things... Full disclosure: I do not study vocal communication specifically, nor do I study vocal development. Infants seem to express a pretty full vocal range at an early age. Within a few months they can make all of the typical chimp sounds: grunts and barks and hoots, etc. but, from what I’ve seen, they don’t consistently apply them appropriately for at least a year. For instance, consider pant-grunts: Pant-grunts are a submissive vocalization that subordinate chimps make when approaching or being approached by dominant individuals. Thus the appropriate context for pant-grunting would be when chimps of unequal rank come into close proximity. Very young chimps begin pant-grunting after a few weeks. At first, they seem to sort of mirror their mothers, pant-grunting whenever mom does. But then, after a few months, infant pant-grunts seem to disarticulate from this association and infants begin to pant grunt at everything that moves within 10m of them whether their mother is grunting or not. After a year to 18 months, infants have a much better handle on their communication skills and generally are able to restrict their pant-grunting to appropriate contexts. By Leakey’s age (about 3 years old) they’ve got things nailed down.
What kinds of vocalizations do Lobo and Leakey make at these ages? Do they communicate directly with one another much, or is it generally directed at their mother?Both infants communicate directly with their mom more often than they communicate with other group members, but I don’t think that I’ve ever noticed either baby using their mother as a conduit for communication with their sibling. Like other mothers, sometimes Lia will interrupt another individual’s attempt to make direct contact with Lobo. For instance if Leakey wishes to play with Lobo but Lia doesn’t want her to Lia might physically prevent Leakey from touching Lobo or make a vocalization that interrupts Leakey’s advances. That said, for the most part, in my experience chimps communicate directly to each other rather than through third parties.  Chimpanzee infants are difficult to sex just by looking at their faces. Can you tell whether these two are male or female? When do you think mothers become aware of their infant’s sex?Pretty immediately. From day one moms spend a lot of time grooming and inspecting every inch of their infants (see below). And despite that there aren't obvious differences in the pelage or facial structure of male and female infants, It isn’t too difficult to determine sex in them. Even though males aren’t born with very conspicuous testes, their penises are pink against their black hair and are easy to see (see below). We can even sex infants in the first few days as long as we can get the right angle on things.
Have you found that mothers respond differently to their infants depending on infant sex?Great question. Let me analyze all this data and get back to you ;) There isn't too much evidence (at the moment) that mamma chimps treat sons differently than they treat daughters, but it can be a hard thing to study, especially in chimps with long inter-birth-intervals. At Kanyawara, there were no cross-sectional differences in the ways that mothers ranged with male versus female infants (Otali et al. 2006). However, There is a paper from the research team at Gombe have reported that individual mothers range differently when they have infant sons versus infant daughters (Murray et al 2014). One of my hopes for this dissertation is to use my data to reexamine ranging patterns and determine if ranging over the last four years is consistent with either or both of these ideas.  One of my favorite shots from the past few months: Wangari relaxing with her sleeping babe while her older two play in the background. What could be better?  pluh-sen-toe-PHAY-gee: postpartum consumption of the mammalian placenta, or afterbirthA brief overview of what I’ve been able to find about placentophagy: While maternal placentophagy is relatively common across mammals, evidence of other, non-mother, placental consumption seems pretty scant. I have not been able to find a previously documented case of non-mother group members consuming placenta in chimps. However, as so many other things, I’m sure that if I’ve seen it, someone else must have. I happily, and eagerly, invite anyone and everyone to send me any evidence that you may have- whether from personal experience or someone else’s documented account. Witnessing a live birth is extremely rare among wild chimpanzees (but check out this article from Nature about the similarities between human and chimpanzee birthing processes). Because of this, the data is scant, but I’ve gathered what I can about placentophagy at other sites. Researchers at Bossou, Guinea recently reported a their second case of researchers witnessing a live birth (Fujisawa et al. 2016). In that instance, the mother consumed the placenta and, although others gathered round and inspected the baby, she did not share it with any other individuals. I learned through personal communication that there was a case of one individual female at Gombe carrying her placenta around for days after parturition until it shriveled up and turned black. While several other individuals approached her during this time and were clearly very interested in the placenta, no one other than the mother ever ate any part of it. And then there's Kanyawara. Luckily, I've been surrounded by field assistants and PI's over the last few weeks and have been able to ask a million questions about their previous experiences here. I confirmed one report of a female burying her placenta after birth. In a second case, a different female ate her own placenta after birth. However, in most cases that we've had an observer in the right place at the right time, the placenta was never seen. This certainly does not preclude placentophagy as mothers generally build large day nests and give birth in those nests, only emerging hours after parturition to rejoin the group (Gombe: Goodall 1980, Kanyawara: unpublished data). We cannot enter such nests to check for the presence/absence of the placenta so it is impossible to comment on whether they were consumed. Further, I have seen older siblings (including this particular placentophile, Gola) entering birthing nests on multiple occasions both while mothers and newborns are still in them, and after they have left. This may provide those siblings with the opportunity to access and consume their mother’s placentas but, again, is very difficult to confirm or refute due to nesting conditions. So again: calling all field biologists! Send more information! You can even use the comments section of this blog post to add your experience or any cases that you’ve read about or even just to name some people that I should ask about it! I am really interested to know if what we saw was truly “unique” or just under-reported! Sources:
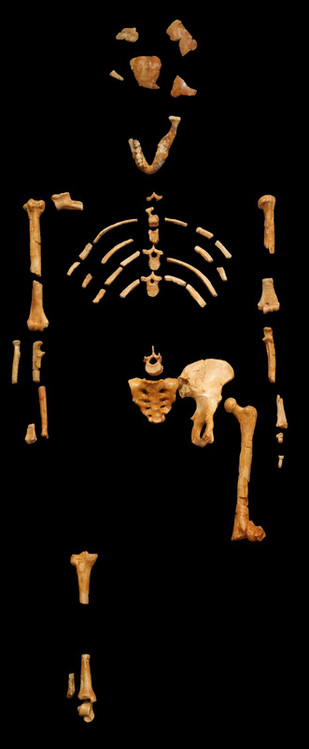
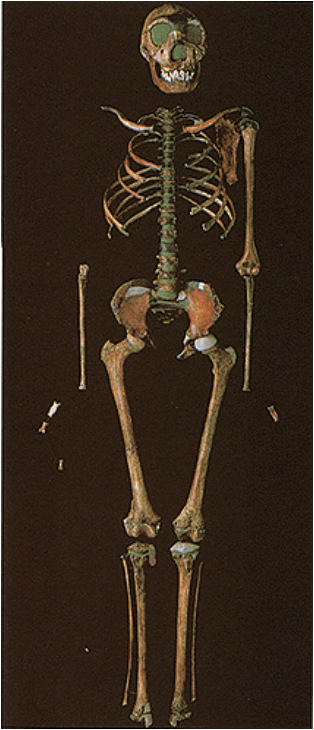
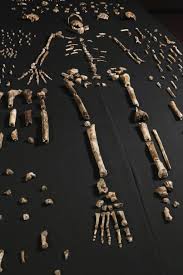
1. Fujisawa, M., Hockings, K. J., Soumah, A. G., & Matsuzawa, T. (2016). Placentophagy in wild chimpanzees (Pan troglodytes verus) at Bossou, Guinea. Primates, 57(2), 175-180. 2. Goodall, J., & Athumani, J. (1980). An observed birth in a free-living chimpanzee (Pan troglodytes schweinfurthii) in Gombe National Park, Tanzania. Primates, 21(4), 545-549.  I’ve been working on this post, with my best and brilliant friend, Marian Hamilton, for months now. We keep waiting for the Naledi news calm down enough that we can start to summarize what's been going on, but every time we’re ready to pull the trigger, something else hits the newsstands. Right now its shenanigans about whether the specimens could have been murdered, sacrificed, or some other form of cultural violence led to their deaths (more on that in subsequent posts). Anyway, we've decided that this piece is better suited as pieces and so we split things up a bit. Part One of Have you met Naledi? is called Naledi's 1,500 Parts. This piece summarizes news reports, science blogs, and published manuscripts describing the Dinaledi site and physical specimens that comprise Homo naledi. We will discuss the descriptive observations and measurements that we have so far and try to bring all of the pieces together (heavily referenced and linked to original sources, of course) with a little more emphasis on the science than most pop-science and media outlets and a little less intense detail than the original manuscripts from the team. The point of this section is to help situate the evidence in the context of our current understanding of human evolution. Part Two (or possibly parts Two+) will be Naledi in the Crux of Controversy and will focus on a number of controversies that swirl around H. naledi. First, we’ll discuss the more controversial ideas from Berger’s team and others about how these specimens made it to the cave where they were discovered. Second, we’ll delve into the bigger debates about how we, evolutionary and paleoanthropologists in particular, do our science and report our findings. Third, we’ll talk about Berger’s hiring decisions, international reactions to a largely female excavation team, and whether this represents a case study of sexism in science.  image credit: thesecondcity.com 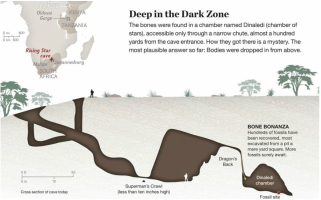 The location of the Rising Star Cave System and illustration of the cave itself. image credit: National Geographic The location of the Rising Star Cave System and illustration of the cave itself. image credit: National Geographic Part One: Naledi’s 1,500 Parts Keeping up with current research and new discoveries can be hard, but sometimes we get those publication gems that make it easy- piercing through to the front page and attracting hoards of media attention. Even those of you who do not obsessively follow new findings in paleoanthropology have probably heard of Homo naledi, the newest member of the genus Homo and our extended ancestral family. At the very least, I bet you caught the initial excited press releases and immediately following sharp edges of media controversy surrounding this assemblage (more on that in subsequent posts). With all this buzz, and the smattering of drama, it can be hard to cut through everything and keep all the information straight. The purpose of this post is to gather scholarly bits and pop-sci pieces from around the web and distill them into some solid science knowledge, Buzzfeed style: 8 (sets of) things to know about Homo naledi: 1. H. naledi comes from South Africa. The taxonomic name, Homo naledi, derives from the Dinaledi chamber, or “Chamber of Stars,” where the specimens were discovered. The chamber is part of the Rising Star Cave System near Johannesburg, South Africa and is connected to the rest of the system by a single chute called the ! The system has been explored for at least 50 years and people had even entered the Dinaledi chamber multiple times prior to the 2013 discovery of this assemblage. 2. This is a MASSIVE set of remains! The Dinaledi Chamber assemblage includes more than 1500 bone fragments from at least 15 individuals that range in age from infant to geriatric! This single site has unearthed more bones representing a single species than almost any other ancestral hominin site and is the largest, richest assemblage every discovered on the continent of Africa. In general, Most paleoanthropological discoveries are limited to single bone or tooth, or perhaps a few clustered together. The best known and most charismatic hominin finds - Lucy the australopithecine from Ethiopia, the Nariokatome Boy (we've included pictures of each skeleton below), the Homo erectus from Kenya - are fairly complete individuals, but were discovered in isolation from others of their species. This makes it very hard for researchers to understand how varied the species might have been in terms of size, dimensions, or cranial size. Imagine millions of years from now if the only modern human to fossilize and be discovered was the 7.5-foot tall NBA star Yao Ming. What would future paleoanthropologists conclude about humans from 2015? Of course this is a very extreme example, but it illustrates the broader point: the more individuals we find of a given species, especially all together in the same place, the better understanding we have of what ‘normal’ might have been for that species. That is what makes the Dinaledi Chamber so valuable!
Two of the other most fruitful hominin excavation sites, Sima de los Huesos in Spain and the Dminisi site in Georgia (the country of Georgia, of course) can help us put this assemblage in perspective. Since 1976, Sima, the largest excavation of hominin fossils to date, has yielded 28 individuals and 6,500 bone fragments. Dmanisi, also regarded as one of the most productive hominin sites, has 5 excavated individuals. The Rising Star expedition has yielded 15 individuals over two to three years at Denaledi- already tripling the number of individuals at Dmanisi. Compared to Sima, the site has recovered over half the number of individuals and a quarter of the number of bone fragments, but in about ONE 20TH OF THE TIME. Can you see why people (including Marian and I) are so excited about all this?!
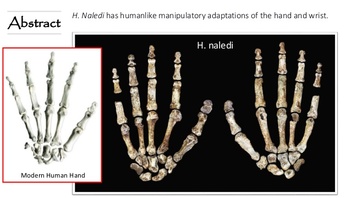
3. This is (probably) a new leaf, or species, on the tree of human evolution. The experts are arguing about this because that’s what they do, but it looks like we have a new species of ancient hominin. H. naledi is represented by multiple (nearly) complete skeletons. Morphologically, they are a mash-up of hominin and australopithecine traits, which sends us some mixed messages. This is to say that they don’t fit in with any other previously discovered hominin or australopithecine. They are, “Weird as hell,” to quote paleoanthropologist Fred Grine from Stony Brook University. “How so?” you ask. Well, let us tell you…This is about to get a little nitty gritty- so hold tight! 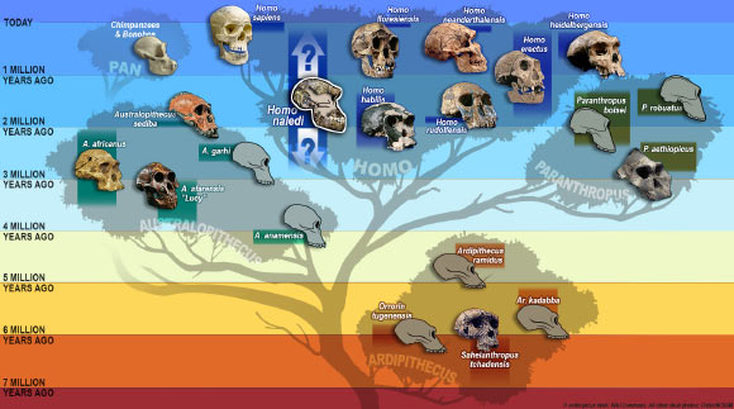 Our current ideas about the human evolutionary tree and where Naledi might fit in. Image credit: S.V. Maderis, University of Wisconsin, Madison 4. H. naledi’s mixed morphology: some parts look more like us…
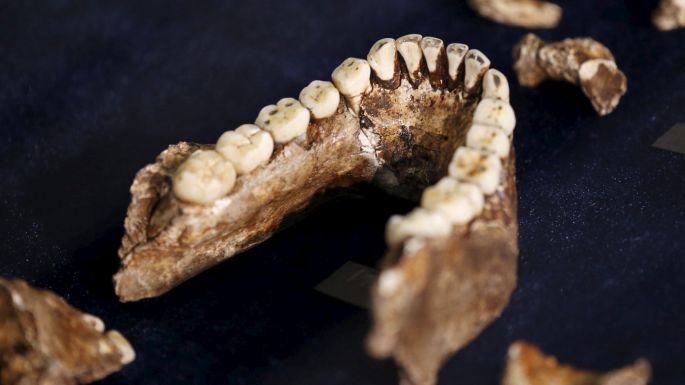 Image Credit: Siphiwe Sibeko, Reuters 5. ....and other parts look more like australopithecines.
 Seriously, if you haven't checked out the National Geographic coverage on this, you should. Their infographics are pretty damned great. Image credit: National Geographic 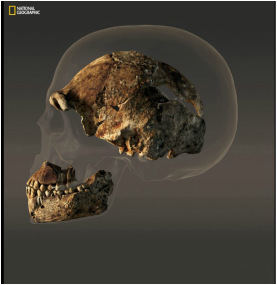 A reconstructed H. naledi skull laid over an average-sized modern human. Image credit: National Geographic A reconstructed H. naledi skull laid over an average-sized modern human. Image credit: National Geographic 6. Ok, fine, but what about brain size? Ah, yes, the other diagnostic marker of a “true” hominin… H. naledi had a brain size of about 560cc, which is less than half the brain volume of the average human (averaging 1350cc) and barely larger than chimpanzees (275-500cc). Gracile australopithecines, like Lucy, fall just below H. naledi between 420 and 450cc, and robust australopithecines, like A.boisei, are just about the same size, averaging around 530cc. By the time we get to Homo, on the other hand, cranial volume surpassed H. naledi. The earliest member of the genus Homo (2.1-1.5 mya), H. habilis, overlaps with H. naledi at 510-660cc1 on the lower end, but by H. erectus (also classified as H. ergaster), cranial capacity is up to 850cc and growing. However, even though the size of the brain is more like earlier australopithecines than Homo, the shape of the crania is much more similar to Homo than earlier hominins. 7. We don't know how old H. naledi is...yet. Dating cave-people is hard! While caves are awesome at preserving bones and fossil assemblages, accurately dating bones that have been recovered from a cave - particularly this cave - is prohibitively difficult. Why? Well let’s take a look our current toolkit for dating fossils:
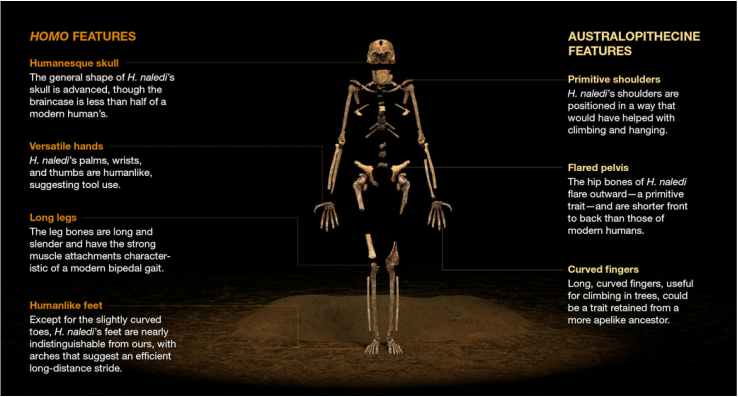
 Image Credit: Curtis, Eye Witness News 8. “But, wait a second, didn’t I just see a report that Naledi was 2-3 million years old?” Yes, impressively well-informed reader, you did. A recent study by Thackeray compared a composite measure of cranial morphology between H. naledi and other human ancestors, placing H. naledi between two other hominins: H. rudolfensis and H. erectus. The maximum recorded age for H. rudolfensis is about 2.5 million years and H. erectus is considered to be about 1.5 million years old. As such, the author concluded that the H. naledi branch must be somewhere on the order of 2 million years old. However, this method of comparing the shape and size of H. naledi bones versus other, more confidently-dated specimens, is kind of sketchy. Now, this is a touchy subject among paleoanthropologists and we’re happy to discuss this in depth if anyone is interested, but several studies have shown that phylogenetic trees based on morphology do not faithfully recapitulate genetic relationships. In other words, it isn’t a great method for determining true genetic relation or the position of the branches on an evolutionary tree. Given the mosaic nature of H. naledi’s skeletal features, this type of dating may be particularly problematic. Think about it this way: when the mosaic of traits exhibited by a single set of skeletons is representative of two different genuses that span a few million years, which features can you choose as the “most” relevant? The classic approach is to look for the most "derived" traits, or the traits that are present in more recent ancestors, but not older ones. However, its pretty easy to see how this sort of classification can get real circular real fast. And that's why we’re on the side of keeping this issue open for discussion.  Image credit: Dog on the Moon for the Guardian Image credit: Dog on the Moon for the Guardian Ok, so obviously this cartoon is ridiculous- I mean, the hominins are dogs. But this cartoonist (Dog on the Moon) drew up a pretty hilarious piece depicting all the to-do about Naledi. Check out the full cartoon, but only if you promise to remember that it's satire. So to recap this hodgepodge of a hominin: Homo naledi had human-like feet and long, strongly muscled legs adapted for striding, walking, and perhaps even running, but a shoulder joint and long, curved fingers that look more like a species spending time in the trees. They had flaring, australopithecine-like hips and a wide pyramid-shaped ribcage, but Homo-like vertebrae. Their teeth were small like ours but simple in their shape and structure; their brain size was small like earlier species, but the shape of the cranium shared many traits with other members of the genus Homo. The real question is: what are we to make of all this? First, the mixed morphology of Naledi flies in the face of popular notion that there was a “human adaptive package” - a suite of traits that arose all together that shifted us from not-human to human. Typically, these traits include larger brain size, hand adaptations for increased dexterity, smaller dentition and smaller guts, and adaptations for efficient long-distance walking and running. H. naledi makes it pretty clear that these traits did not arise all together, and even throw into question the logical order in which such traits would have arisen. For example, most people assume that large brain size was a prerequisite for sophisticated tool use, and then anatomical adaptations for fine motor skills and manipulation would have come afterwards. With their human-like hands but australopithecine-like brains, H. naledi looks from the past and suggests, “Maybe not…” Second, no matter how old these specimens are, H. naledi is an incredible find that will revise the story of human evolution. If they preceded humans we might have identified a direct ancestor to our own species. If they lived alongside early Homo, that could have profound implications for the “bushiness” of the hominin branch of the primate tree. However, our ability to fit this evidence into the context of our evolutionary history is severely limited until we find a decent method for dating these specimens. No matter what—this is some very exciting science! Stay tuned for more. Please note: comments and criticisms for this post are welcome, as well as suggestions for future posts. Our goal is to help evolutionary anthropology and science in general reach a wider audience and are always excited for opportunities to do a better job!! Many of the sources used to write this piece are linked directly to the text. Here is a list of those sources plus some additional resources for any and all of those interested: Scientific articles: 1. Berger, L. R., Hawks, J., de Ruiter, D. J., Churchill, S. E., Schmid, P., Delezene, L. K., ... & Skinner, M. M. (2015). Homo naledi, a new species of the genus Homo from the Dinaledi Chamber, South Africa. Elife, 4, e09560. (And associated slideshow: http://www.slideshare.net/Lor342/homo-naledi-highlights-from-the-paper-published-on-elife) 2. Gibbons, A. (2015). New human species discovered. Science, 349(6253), 1149-1150. 3. Harcourt-Smith, W. E. H., Throckmorton, Z., Congdon, K. A., Zipfel, B., Deane, A. S., Drapeau, M. S. M., ... & DeSilva, J. M. (2015). The foot of Homo naledi. Nature communications, 6. 4. Kivell, T. L., Deane, A. S., Tocheri, M. W., Orr, C. M., Schmid, P., Hawks, J., ... & Churchill, S. E. (2015). The hand of Homo naledi. Nature communications, 6. 5. Stringer, C. (2015). The many mysteries of Homo naledi. eLife, 4, e10627. 6. Thackeray, J. F. (2015). Estimating the age and affinities of Homo naledi.South African Journal of Science, 111(11 & 12), 1-2. Other sources: Team Leader, Lee Berger's website: http://ewn.co.za/Features/Naledi Paleoanthropologist and team member John Hawke's Blog: http://johnhawks.net/ The Expedition Blog: http://voices.nationalgeographic.com/blog/rising-star-expedition/ National Geographic Coverage: http://news.nationalgeographic.com/2015/09/150910-human-evolution-change/ Other news releases: http://theconversation.com/homo-naledi-may-be-two-million-years-old-give-or-take-50416 http://www.sciencemag.org/content/349/6253/1149.full https://fsp.trinity.duke.edu/projects/new-fossil-discovery-redefines-what-it-means-be-human Other blogs about Naledi: http://ncse.com/blog/2015/10/cave-homo-naledi-textbook-example-how-to-do-science-0016693 For some words about Naledi from Berger's own mouth, check out this video!
|
Kris SabbiThis blog is a forum share my personal experiences as a field researcher and traveler. Categories
All
Blog Archives
June 2022
By Category
All
|















 RSS Feed
RSS Feed
